何謂『禁色』耶?既然得『禁』,特明示也。表示很可能發生乎?借來翻譯『無法看見』之顏色,頗懸疑焉!
Impossible color
Impossible colors or forbidden colors are supposed colors that cannot be perceived in normal seeing of light that is a combination of various intensities of the various frequencies of visible light, but are reported to be seen in special circumstances.
![]()
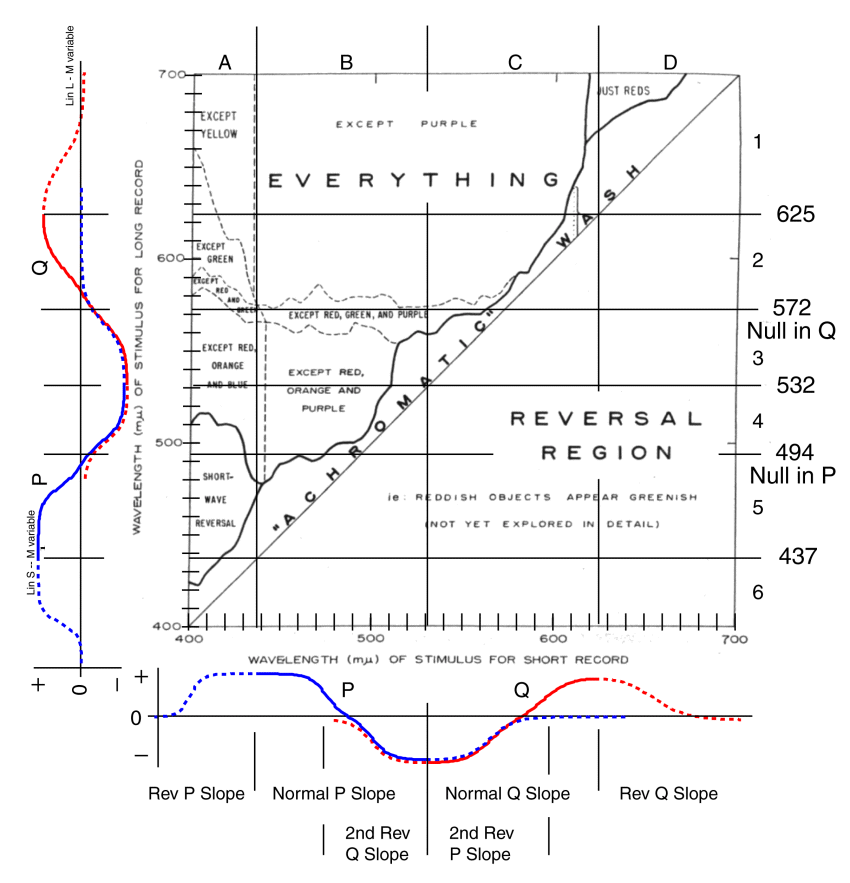
The human eye’s red-to-green and blue-to-yellow values of each one-wavelength visible color
Types
These impossible colors are of two types:
- Colors that would be seen if the output strengths of the human eye retina‘s three types of cone cell (red, green, blue) could be set to values which cannot be produced by exposing the eye in normal seeing conditions to any possible combination of strengths of the frequencies of visible light.
- Colors that cannot be seen directly from any combination of retina signal output from one place in one eye, but can be generated in the brain’s visual cortex by mixing color signals from the two eyes, or from more than one part of the same eye. Examples of these colors are bluish-yellow and reddish-green.[1] Those colors that appear to be similar to, for example, both red and green, or to both yellow and blue. (This does not mean the result of mixing paints of those two colors in painting, or the result of mixing lights of those two colors on a screen.)
Opponent process
The color opponent process is a color theory that states that the human visual system interprets information about color by processing signals from cone and rod cells in an antagonistic manner. The three types of cone cells have some overlap in the wavelengths of light to which they respond, so it is more efficient for the visual system to record differences between the responses of cones, rather than each type of cone’s individual response. The opponent color theory suggests that there are three opponent channels:
- Red versus green.
- Blue versus yellow
- Black versus white (this is achromatic and detects light-dark variation, or luminance).
Responses to one color of an opponent channel are antagonistic to those to the other color, and signals output from a place on the retina can contain one or the other but not both, for each opponent pair.
假使那『不可能』的色彩可以看見,就得知道『什麼所禁』哩!!似乎過去『科技變遷』,逐漸改變著『觀察者』之『主、客觀性』的分野︰
相對論將『觀察者』帶入物理,改變了『量測』的基本觀念。雖然無限精準的『測量』即使作不到,尚且還可以想像。但是量子力學把『量測』的『測不準』原理放進物理,就是說連想像『粒子』的『軌跡』在原理上都不『允許』!!量子力學是使用著『運算子』operator 的語言來描寫微小粒子之『事件概率』的『波函數』。那『測不準原理』是什麼呢?所謂『經典物理學』classic physics 對一個『物體』運動軌跡的描述是由它的『位置』和『速度』或說『動量』所確定的,一九二七年德國的維爾納‧海森堡 Werner Heisenberg 卻講任何『量子系統』之『量測』必為如下的關係式所制約︰


這並不是因為觀察者的量測,影響了系統── 比方說用粗大的溫度計量一小杯水的溫度 ──所導致的『觀察者效應』,而是宇宙的本質如此。所以即使是想像一個箱子裡的『電子軌跡』都沒有『旨趣』,你不量測它想說它是『波』或者是『粒子』之象純屬『無謂』。這引發一些物理學大方家不滿,認為量子力學根本尚未『完備』。就像發展完成量子力學『波動方程式』的埃爾溫‧薛丁格,他卻也是提出一個稱之為『薛丁格貓 』之想像實驗的人,用以表達目前量子力學之『哥本哈根詮釋』所必須思考的嚴峻性矛盾問題︰
薛丁格是如此描述這個實驗的:
實 驗者甚至可以設置出相當荒謬的案例來。把一隻貓關在一個封閉的鐵容器裡面,並且裝置以下儀器(注意必須保固 這儀器不被容器中的貓直接干擾):在一台蓋革計數器內置入極少量放射性物質,由於物質的數量極少,在一小時內,這個放射性物質至少有一個原子衰變的機率為 50%,它沒有任何原子衰變的機率也同樣為50%;假若衰變事件發生了,則蓋革計數管會放電,通過繼電器啟動一個榔頭,榔頭會打破裝有氰化氫的燒瓶。經過 一小時以後,假若沒有發生衰變事件,則貓仍舊存活;否則發生衰變,這套機構被觸發,氰化氫揮發,導致貓隨即死亡。用以描述整個事件的波函數竟然表達出了活 貓與死貓各半糾合在一起的狀態。
類似這典型案例的眾多案例裏,原本只局限於原子領域的不明確性被以一種巧妙的機制變為宏觀不明確性,只有通過打開這個箱子來直接觀察才能解除這樣的不明確 性。它使得我們難以如此天真地接受採用這種籠統的模型來正確代表實體的量子特性。就其本身的意義而言,它不會蘊含任何不清楚或矛盾的涵義。但是,在一張搖 晃或失焦的圖片與雲堆霧層的快照之間,實則有很大的不同之處。
不僅如此,在量子系統中,假使兩個粒子在經過短時間彼此間耦合之後,儘管將這兩個粒子分隔很遠的一段距離,量測其中任何一個粒子,會不可避免地影響到另外一個粒子的度量性質,彷彿有隔空的傳心術一般,這種關聯現象稱之為『量子糾纏 』quantum entanglement 。
當初愛因斯坦,波多爾斯基和羅森三人提出 ── Albert Einstein, Boris Podolsky and Nathan Rosen EPR paradox ── 這個悖論的 目的是想用,沒有任何『物理訊息』── 量子糾纏也該不行 ──的傳播能夠超過『光速』,來證明量子力學的不完備性。但是多次重複所做的實驗已經證實量子糾纏的這個論點,也就是說,量子糾纏的速度確實比光速還要 快。最近完成的一項實驗顯示,量子糾纏的作用速度至少比光速快上萬倍,這還只是速度的下限,因為根據量子理論,測量效應是瞬時的。
人類打開了大自然的『天書』,讀取了其中『幾頁』,到底該如何『理解』進而能『詮釋』它呢?許多跡象顯示現今的人們多半只愛談『應用』,至於到底『電子』是存在的嗎?或只是為著理論的『方便』所作的『虛構』的呢?假使它果存在,又為什麼時而是『粒子』時而是『波』的呢?……就留與其人了!!
─── 摘自《測不準原理》
莫非『禁色』預告『色彩典範』將要變遷了嗎??

The Physiological Explanation ofLand’s Retinex Theory
Prepared by James T. Fulton
Introduction
In 1958, Dr Edwin H. Land, founder and principle stockholder in the Polaroid Corporation, gave a series of demonstrations of unusual visual effects that he asserted could not be explained with the Theory of Color Vision accepted at that time. While a highly respected business man and scientist, he was also a superb showman on stage. However, he was an experimentalist of the Thomas A. Edison school. At the time he offered no theoretical explanation of his findings but merely undermined the generally excepted theory.
In his prepared remarks and the first paragraph of his first published paper1, Land asserted that “We have come to the conclusion that the classical laws of color mixing conceal great basic laws of color vision. There is a discrepancy between the conclusions that one would reach on the basis of the standard theory of color mixing and the results we obtain in studying total images.” His reference is to the conventional wisdom that vision was/is based on additive color mixing. “This departure from what we expect on the basis of colorimetry is not a small effect but is complete, and we conclude that the factors in color vision hitherto regarded as determinative are significant only in a certain special case. What has misled us all is the accidental universality of this special case.”
In his second paper2, Land summarized the observed phenomena in a single figure that has been modified here by the addition of more finely divided wavelength scales. He concluded that the perception of color “depends on a ratio of ratios; namely, as numerator, the amount of a long-wave stimulus at a point as compared with the amount that might be there; and, as a denominator, the amount of a shorter wave stimulus at that point as compared with the amount that might be there.” Although, Land was approaching the actual case, he remained far from a precise definition of the situation (and the terms he used lacked precision in the context of today).
Because of the lack of rigorous scientific controls in the course of his public audience-based demonstrations and his obvious skills as a showman, his results were not received well by the scientific community to which he was a prominent member. “Although generally regarded as fascinating research by the physicists, and the research community, these observations irritated, if not inflamed, the color vision community4.” Many considered the experiments as containing subtle events that introduced changes in adaptation and other phenomena not unlike those achieved in the typical magic act in order to control the perception of the audience.
The third paper in the series, promised in the closing of the second, never appeared. However, a paper defining an initial version of the Retinex Theory of Color Vision, coauthored with his long-time colleague, did appear in 1971, twelve years later3. The theory was largely conceptual but did assert it applied to a system of three spectrally narrowband photoreceptors. The concept of a broadband rod was not included in the paper. The discussion was based on a three projector configuration using three fixed filters with spectral widths of 50-100 nm. It did not refer to or offer a rigorous explanation of the experiments in the second paper.
Land & McCann defined the term retinex to describe each of the spectral subsystems of the complete retinal system applicable to vision. They describe a system of three spectrally separate subsystems extending from the retina to the cortex. They assert the spectral images are not mixed but are compared. Beyond that assertion, they did not discuss the architecture or the specific circuitry of the neurological system. Looking back, the context of their work makes it clear they were thinking in terms of the linear operation of the visual system as appropriate to the lower photopic and mesotopic regime in all three papers.
The Retinex Theory as originally conceived is incompatible with the current zone models of vision where significant spectral signal differencing occurs in the retina. The zone model defined in this work is presented in the Visual Block Diagram for humans The validity of the proposed zone model includes its ability to describe the results of the Land experiments in detail.
A more detailed description of the zone model of this theory is shown in the Overall Block Diagram.
The title Retinex Theory has been transformed in the subsequent technical literature so that the title is also used to describe methods, largely computer algorithms, to improve the contrast performance of imagery containing large dynamic ranges (basically improving the visibility of detail obscured in shadowed areas of scenes). Attempts have also been made to show the Retinex Theory underlies the phenomenon of color constancy. However, the conceptual level of the theory has not led to a definitive explanation of color constancy5.
The Electrolytic Theory of the Visual Process (a title used on this page to avoid confusion due to the alternate title, Tetrachromatic Theory of Biological Vision) presented in this work provides a rigorous explanation of Dr. Land’s experiments and provides additional insights into those experiments that can be experimentally verified. This will be demonstrated below. This paper does not concern itself with color constancy or contrast enhancement of scene details. A different concept and architecture of color constancy is offered in Chapter 7.
Subsequent to the arrival of the INTERNET, something of a cottage industry has arisen attempting to develop algorithms to perform contrast enhancement based on Land’s findings. While consisting of interesting mathematical manipulations in computer controlled spaces, most have not proven robust. Their applicability to biological vision has been distinctly limited.
※註【機制關聯性】
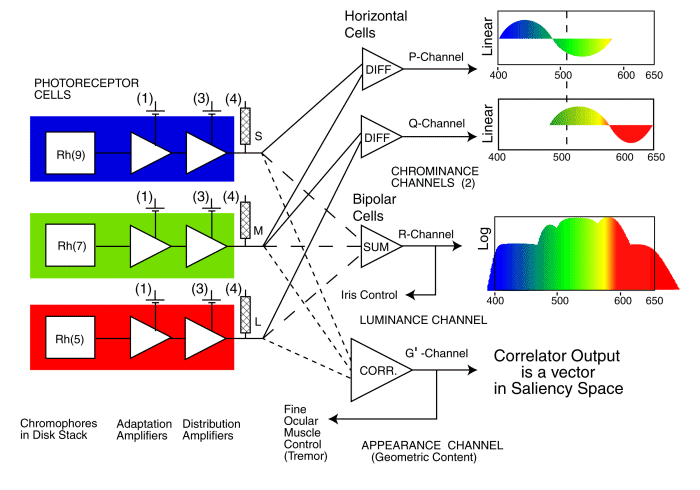
SIGNALING BLOCK DIAGRAM OF THE VISUAL SYSTEM IN HUMANS
INTRODUCTION
The signaling architecture of the visual system used in long wavelength trichromats (humans and other large terrestrial mammals) is shown in the following figure. This figure emphasizes the three distinct signaling paths found in vision (the P-, Q- and R-channels) as well as the unique correlation channel (labeled the G’-channel) found in only the highest primates (hominoidea) and used by the analytical mode of vision for reading and other functions requiring maximum acuity and perception of detail.
Opponent process
The color opponent process is a color theory that states that the human visual system interprets information about color by processing signals from cones and rods in an antagonistic manner. The three types of cones (L for long, M for medium and S for short) have some overlap in the wavelengths of light to which they respond, so it is more efficient for the visual system to record differences between the responses of cones, rather than each type of cone’s individual response. The opponent color theory suggests that there are three opponent channels: red versus green, blue versus yellow, and black versus white (the last type is achromatic and detects light-dark variation, or luminance).[1] Responses to one color of an opponent channel are antagonistic to those to the other color. That is, opposite opponent colors are never perceived together – there is no “greenish red” or “yellowish blue”.
While the trichromatic theory defines the way the retina of the eye allows the visual system to detect color with three types of cones, the opponent process theory accounts for mechanisms that receive and process information from cones. Though the trichromatic and opponent processes theories were initially thought to be at odds, it later came to be understood that the mechanisms responsible for the opponent process receive signals from the three types of cones and process them at a more complex level.[2]
Besides the cones, which detect light entering the eye, the biological basis of the opponent theory involves two other types of cells: bipolar cells, and ganglion cells. Information from the cones is passed to the bipolar cells in the retina, which may be the cells in the opponent process that transform the information from cones. The information is then passed to ganglion cells, of which there are two major classes: magnocellular, or large-cell layers, and parvocellular, or small-cell layers. Parvocellular cells, or P cells, handle the majority of information about color, and fall into two groups: one that processes information about differences between firing of L and M cones, and one that processes differences between S cones and a combined signal from both L and M cones. The first subtype of cells are responsible for processing red–green differences, and the second process blue–yellow differences. P cells also transmit information about intensity of light (how much of it there is) due to their receptive fields.[citation needed]

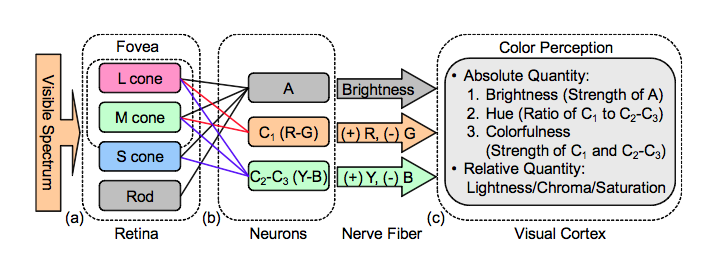
Diagram of the opponent process
欲知是否能夠解釋『夢中色彩』如何形成!☆真能感受既『紅』又『綠』之『蘋果』嗎?☆
還請讀巨著矣☆
PROCESSES IN BIOLOGICAL VISION
by JAMES T. FULTON
This site presents the new, and COMPREHENSIVE
Last update: 1 November 2016 Activa™: See Citation Page
In support of the above,
ARCHITECTURE OF ALL VISION IS TETRACHROMATIC.
Although traditionally called trichromats, it is shown that
HUMANS ARE BLOCKED TETRACHROMATS
For a quick proof of this fact, click here
MULTI-THREAD and MULTI-STAGE ZONE THEORY
The ELECTROLYTIC THEORY presents the first description of the
ACTIVA, the ACTIVE SEMICONDUCTOR KERNAL of EVERY NEURON
The THEORY presents the first
COMPLETE CIRCUIT DIAGRAM from RETINA TO CORTEX.
Many of the most subtle mechanisms associated with the visual modality developed within the context of this theory have been confirmed during 2016. Sabesam et al., working at the University of California, Berkeley assembled a very sophisticated optical coherent tomography (OCT) test set augmented with adaptive (active) optics and tremor cancelling features. This test set allowed them to stimulate individual sensory neuron photoreceptors of the human retina in-vivo. With this capability, they were able to demonstrate that the M – and L–channel photoreceptors mediated the intensity signaling R–channel of the visual modality as well as mediated their individual M – and L–signaling channels. They accomplished this psychophysical experiment by stimulating individual photoreceptors and interrogating the fully aware subject as to their perceived response (Section 3.2.3.6).